Eggs Are Produced Cyclically and Sperm Are Produced Continuously Question
In most species, there are just two types of gamete, and they are radically different. The egg is among the largest cells in an organism, while the sperm (spermatozoon, plural spermatozoa) is often the smallest. The egg and the sperm are optimized in opposite ways for the propagation of the genes they carry. The egg is nonmotile and aids the survival of the maternal genes by providing large stocks of raw materials for growth and development, together with an effective protective wrapping. The sperm, by contrast, is optimized to propagate the paternal genes by exploiting this maternal investment: it is usually highly motile and streamlined for speed and efficiency in the task of fertilization. Competition between sperm is fierce, and the vast majority fail in their mission: of the billions of sperm released during the reproductive life of a human male, only a few ever manage to fertilize an egg.
Sperm Are Highly Adapted for Delivering Their DNA to an Egg
Typical sperm are "stripped-down" cells, equipped with a strong flagellum to propel them through an aqueous medium but unencumbered by cytoplasmic organelles such as ribosomes, endoplasmic reticulum, or Golgi apparatus, which are unnecessary for the task of delivering the DNA to the egg. Sperm, however, contain many mitochondria strategically placed where they can most efficiently power the flagellum. Sperm usually consist of two morphologically and functionally distinct regions enclosed by a single plasma membrane: the tail, which propels the sperm to the egg and helps it to burrow through the egg coat, and the head, which contains a condensed haploid nucleus (Figure 20-25). The DNA in the nucleus is extremely tightly packed, so that its volume is minimized for transport, and transcription is shut down. The chromosomes of many sperm have dispensed with the histones of somatic cells and are packed instead with simple, highly positively charged proteins called protamines.
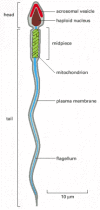
Figure 20-25
A human sperm. It is shown in longitudinal section.
In the head of most animal sperm, closely apposed to the anterior end of the nuclear envelope, is a specialized secretory vesicle called the acrosomal vesicle (see Figure 20-25). This vesicle contains hydrolytic enzymes that may help the sperm to penetrate the egg's outer coat. When a sperm contacts an egg, the contents of the vesicle are released by exocytosis in the so-called acrosome reaction; in some sperm, this reaction also exposes or releases specific proteins that help bind the sperm tightly to the egg coat.
The motile tail of a sperm is a long flagellum, whose central axoneme emanates from a basal body situated just posterior to the nucleus. As described in Chapter 16, the axoneme consists of two central singlet microtubules surrounded by nine evenly spaced microtubule doublets. The flagellum of some sperm (including those of mammals) differs from other flagella in that the usual 9 + 2 pattern of the axoneme is further surrounded by nine outer dense fibers (Figure 20-26). These dense fibers are stiff and noncontractile, and it is not known what role they have in the active bending of the flagellum, which is caused by the sliding of adjacent microtubule doublets past one another. Flagellar movement is driven by dynein motor proteins, which use the energy of ATP hydrolysis to slide the microtubules, as discussed in Chapter 16. The ATP is generated by highly specialized mitochondria in the anterior part of the sperm tail (called the midpiece), where the ATP is needed (see Figures 20-25 and 20-26).
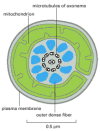
Figure 20-26
Drawing of the midpiece of a mammalian sperm as seen in cross section in an electron microscope. The core of the flagellum is composed of an axoneme surrounded by nine dense fibers. The axoneme consists of two singlet microtubules surrounded by nine microtubule (more...)
Sperm Are Produced Continuously in Most Mammals
In mammals, there are major differences in the way in which eggs are produced (oogenesis) and the way in which sperm are produced (spermatogenesis). In human females, for example, oogonia proliferate only in the fetus, enter meiosis before birth, and become arrested as oocytes in the first meiotic prophase, in which state they may remain for up to 50 years. Individual oocytes mature from this strictly limited stock and are ovulated at intervals, generally one at a time, beginning at puberty. In human males, by contrast, meiosis and spermatogenesis do not begin in the testes until puberty and then go on continuously in the epithelial lining of very long, tightly coiled tubes, called seminiferous tubules. Immature germ cells, called spermatogonia (singular, spermatogonium), are located around the outer edge of these tubes next to the basal lamina, where they proliferate continuously by mitosis. Some of the daughter cells stop proliferating and differentiate into primary spermatocytes. These cells enter the first meiotic prophase, in which their paired homologous chromosomes participate in crossing-over, and then proceed with division I of meiosis to produce two secondary spermatocytes, each containing 22 duplicated autosomal chromosomes and either a duplicated X or a duplicated Y chromosome. The two secondary spermatocytes derived from each primary spermatocyte proceed through meiotic division II to produce four spermatids, each with a haploid number of single chromosomes. These haploid spermatids then undergo morphological differentiation into sperm (Figure 20-27), which escape into the lumen of the seminiferous tubule (Figure 20-28). The sperm subsequently pass into the epididymis, a coiled tube overlying the testis, where they undergo further maturation and are stored.
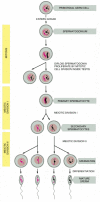
Figure 20-27
The stages of spermatogenesis. Spermatogonia develop from primordial germ cells that migrate into the testis early in embryogenesis. When the animal becomes sexually mature, the spermatogonia begin to proliferate rapidly, generating some progeny that (more...)

Figure 20-28
Highly simplified drawing of a cross section of a seminiferous tubule in a mammalian testis. (A) All of the stages of spermatogenesis shown take place while the developing gametes are in intimate association with Sertoli cells. These large cells extend (more...)
An intriguing feature of spermatogenesis is that the developing male germ cells fail to complete cytoplasmic division (cytokinesis) during mitosis and meiosis. Consequently, large clones of differentiating daughter cells that have descended from one maturing spermatogonium remain connected by cytoplasmic bridges, forming a syncytium (Figure 20-29). The cytoplasmic bridges persist until the very end of sperm differentiation, when individual sperm are released into the tubule lumen. This accounts for the observation that mature sperm arise synchronously in any given area of a seminiferous tubule. But what is the function of the syncytial arrangement?

Figure 20-29
Cytoplasmic bridges in developing sperm cells and their precursors. The progeny of a single maturing spermatogonium remain connected to one another by cytoplasmic bridges throughout their differentiation into mature sperm. For the sake of simplicity, (more...)
Unlike oocytes, sperm undergo most of their differentiation after their nuclei have completed meiosis to become haploid. The presence of cytoplasmic bridges between them, however, means that each developing haploid sperm shares a common cytoplasm with its neighbors. In this way, it can be supplied with all the products of a complete diploid genome. Developing sperm that carry a Y chromosome, for example, can be supplied with essential proteins encoded by genes on the X chromosome. Thus, the diploid genome directs sperm differentiation just as it directs egg differentiation.
Some of the genes that regulate spermatogenesis have been conserved in evolution from flies to humans. The DAZ gene, for example, which encodes an RNA-binding protein and is located on the Y chromosome, is deleted in many infertile men, many of whom cannot make sperm. Two Drosophila genes that are homologous to DAZ are essential for spermatogenesis in the fly. RNA-binding proteins are especially important in spermatogenesis, because many of the genes expressed in the sperm lineage are regulated at the level of RNA translation.
Summary
A sperm is usually a small, compact cell, highly specialized for the task of fertilizing an egg. Whereas in human females the total pool of oocytes is produced before birth, in males new germ cells enter meiosis continually from the time of sexual maturation, with each diploid primary spermatocyte giving rise to four haploid mature sperm. The process of sperm differentiation occurs after meiosis is complete, requiring five weeks in humans. Because the maturing spermatogonia and spermatocytes fail to complete cytokinesis, however, the progeny of a single spermatogonium develop as a large syncytium. Sperm differentiation is therefore directed by the products from both parental chromosomes, even though each nucleus is haploid.


Source: https://www.ncbi.nlm.nih.gov/books/NBK26914/
0 Response to "Eggs Are Produced Cyclically and Sperm Are Produced Continuously Question"
Post a Comment